

Why you should know about Harold Hillman’s work on the living cell.
by dpl and Mia Breeze | Feb 11, 2024
This paper looks at how Harold Hillman’s critique of the living cell impacts our current understanding of deoxyribonucleic acid (DNA), genetics, viruses, genetically modified organisms (GMO), and our understanding of living things.
I have tried to condense and simplify Hillman’s work concerning the living cell as much as possible to make it more accessible to the average reader but given modern science’s tendency to over complicate and obfuscate things, there is a lot to unravel.
As such, if you are able to put aside your modern attention span and invest time in reading this paper, it is likely you will finish knowing with confidence what exactly about these topics is knowable and what is false.
Introduction
In Harold Hillman’s (b. London 1930 – 2016) obituary, we learn from his brother that,
“… Harold Hillman, … was a biological scientist whose research … repeatedly challenged the orthodox scientific community in its interpretations of the effects on cell structures of extracting, dehydrating and staining under the electron microscope.
In the book The Living Cell (1980), which Harold wrote with Peter Sartory, he argued that the technique resulted in fundamental changes to the cells themselves, and was thus unreliable, a view rejected by most of his peers, and for which he paid a high price in career terms.
Harold was uncompromisingly and intellectually honest, dedicated to leading a life based on principle and driven by a strong moral compass that affected everyone he met. … Its manifestation was revealed in every aspect of his life – his family, his humanist philosophy …, his abhorrence of capitalism and his commitment to exposing institutional corruption, the diminution of academic freedom, cover-ups of discreditable research, loss of human rights and abuse of animal welfare…” (Source 1).
When you go through Hillman’s work and interviews, the above seems to be an accurate description of the man. You can read the story of his career and the prices he paid for his intellectual honesty here (Source 2).
Hillman was a rare scientist, in that the focus of his career was not centred around proving or discovering something novel. Instead, he focused on fact checking and critiquing what are considered proven facts which comprised the very foundations of his chosen field of biology. Facts in the sense that they are ideas taught as infallible and verifiable truths by teachers, lecturers, and authors of biology textbooks to this day.
When you read Hillman’s books, The Living Cell (1980) and Certainty and Uncertainty in Biochemical Techniques (1972), you get a good idea of Hillman’s uncompromising intellectual honesty. However, once you have gone through a substantial amount of Hillman’s work you begin to realise that demonstrating intellectual honesty was the underlying theme of all his scientific work, the actual focus of his career.
Intellectual honesty in scientific experiments in any branch requires the methodical and good faith application of the scientific method.
This means the identification of all assumptions including those related to the procedures undertaken to carry out the experiment – i.e. the limitations of any apparatus, preparations and/or procedures followed to carry out the experiment. It also means drawing attention to how those limitations impact the final conclusions drawn from the experiment.
It means undertaking control experiments at appropriate stages of the experiment, which includes accuracy experiments relating to the equipment used. It means publishing the results of those control experiments in every case and especially when they cast doubt on the initial hypothesis. Refer to Hillman, H. (1972). Certainty and Uncertainty in Biochemical Techniques. Surrey University Press, Henley on Thames.
Application by Hillman of the scientific method in the manner described above to the studies of cells and their inner structures, lead Hillman to make some profound criticisms of modern biology.
As stated by Tom Cowan, Hillman’s work demonstrated that many of the foundational facts of biology were very far from facts so much so that the whole field of modern cell biology, the underpinnings of modern medicine, had to be re-evaluated (Source 4). From what I have read of Hillman’s work, Cowan was not exaggerating.
Why should you care about the field of modern cell biology?
Well, because how we think about the cell underpins how we think living entities operate. The cell is considered to be the smallest living thing out of which every living thing is composed – the basic unit of life. Cells are said to run all the processes needed to sustain life, such as protein synthesis. Cells are said to originate only from other cells.
Our DNA, and thus our genes, are said to reside in our cells which are replicated and passed on to every daughter cell formed via division. This is how genes and cells are said to be responsible for our inherited characteristics. Genes are also said to be responsible for a person or animals’ susceptibility to certain diseases.
So, what are these profound criticisms of cell biology?
The majority of Hillman’s work has far reaching implications for modern biology and medicine, but given that this paper is only big enough to cover one aspect of his work in detail, I have tried to choose an aspect which is likely to be the most interesting and have the furthest reaching implications – Hillman’s criticism of the structure of the living cell and how it impacts our understanding of protein synthesis. As for the rest of his work, I hope this paper will pique enough interest that the reader will want to look into Hillman’s other work themselves.
Before diving in it is important to be aware upfront what is meant by the ‘living cell’. When Hillman refers to the ‘living cell’ he is referring to a cell that is still within an alive human or animal (in vivo) and not one that has been extracted from the creature (separated from its life source) and treated in order to be analysed.
By making this distinction Hillman highlights the fact that one of the biggest assumptions made by biologists is that removing tissues or cells from its life source has no impact on the sample or its chemistry. In addition, in instances where the animal has to be killed, that the stress of handling and killing the animal has no effect on the results of the experiment.
Hillman and the structures of the generalised cell
Hillman’s reappraisal of the structure of the living cell began in earnest when he and his colleague, Peter Sartory, came across certain important anomalies between the structure and behaviour of the cell as seen by light microscopy in unfixed tissue, on the one hand, and the structure as deduced from electron microscopy in fixed tissue, on the other.
Light microscope and electron microscope
A light or optical microscope focuses a beam of visible light (photons) through a specimen to produce an image with a resolution of 200-250 nanometres (nm). The image thus formed is composed of photons that have not been stopped and those diffracted by the sample.
That image is then passed through one or two lenses to magnify the image for viewing. The stream of photons may be directed at the specimen at any angle, meaning that a natural incidence of the specimen may be selected as if the specimen being examined was in the examiner’s hand.
A light microscopist can observe living, dead and fixed specimens and has at their disposal a wide range of methods of preparation and examination (bright field, phase contrast etc.) but it is only the observation of living specimens under light microscope that will be referred to in this for paper.
When observing living specimens (unstained and unfixed) the sample is merely placed on a microscope slide prior to being placed in the microscope for viewing.
An electron microscope uses a focused beam of electrons to “image” the specimen at a resolution of 1.5 to 2 nm. The process undertaken to produce an electron micrograph is however far more complicated than that required by light microscopy.
First the tissue is fixed (killed) in formalin, this stops metabolism and enzymes are inhibited. Basically, the tissue is treated to prevent it from decomposing before it is viewed.
Next, because the tissue will not survive being put under a high vacuum and the bombardment of electron and x-irradiation, the tissue must be coated or stained by the deposition of heavy metal salts of osmium, lead, tungsten, manganese etc.
The stained tissue is thereafter dehydrated with increasing concentration of alcohol (which shrinks the tissue). The alcohol is then extracted from the tissue with a fat solvent (propylene oxide), following which the tissue is impregnated with and embedded within an epoxy resin (plastic), and then left for a few days to harden.
Sections or slices one tenth of a millimetre thick, or less, are cut with a special cutting tool called a microtome. Note that the microscopists has no idea of the orientation of the microtome blade in relation to the cell and its inner structures when cutting sections.
These incredibly thin sections are then placed inside of the microscope’s viewing chamber where nearly all the air is pumped out to place the specimen under high vacuum, known as venting.
A beam of electrons ranging form 10 00 volts to 3 000 000 volts is directed at the specimen, the amount of volts depends on the magnification desired. The higher the voltage the higher the magnification.
Those electrons which are not intercepted by the heavy metals pass through and strike a fluorescent screen. The microscopists select a particular field (area) of the screen and zooms in until the features they want to demonstrate are observable. The image can then be enhanced, and photographs are taken, some of which are selected as evidence.
Some argue that the more modern rapid deep-freezing techniques, where samples are frozen at -150 degrees or more, can be carried out without causing dehydration of the cellular fluids as is the case with alcohol.
Hillman says that without the undertaking of proper control experiments this has never been demonstrated. Moreover, he disagrees this could be the case given that when something is frozen to very low temperatures, water comes out of any solution it is in, in the form of ice crystals thereby leaving all other constituents of the solutions dehydrated.
It is also essential to realise that over and above the steps that dehydrate the specimen during preparation, the specimen is further dehydrated when subjected to the bombardment of the electron beam. This is because when the electron beam hits the specimen, it heats up the tissue-metal combination causing the temperature of the specimen to rise by hundreds of degrees.
The generalised cell
Following their investigations, Hillman and Sartory concluded that the commonly accepted model of the cell (the generalised cell) is impossible on geometric and biological grounds, and that those inner structures (also known as organelles) deduced by electron microscope are artefacts (products) of the methods employed to prepare the sample for viewing.
Until the early 1940s, before the use of electron microscope to study cells, it was agreed that all living cells were round or spherical entities consisting of an outer or cell membrane enclosing the sphere, mitochondria, a nucleus, cytoplasm and a nucleus membrane.
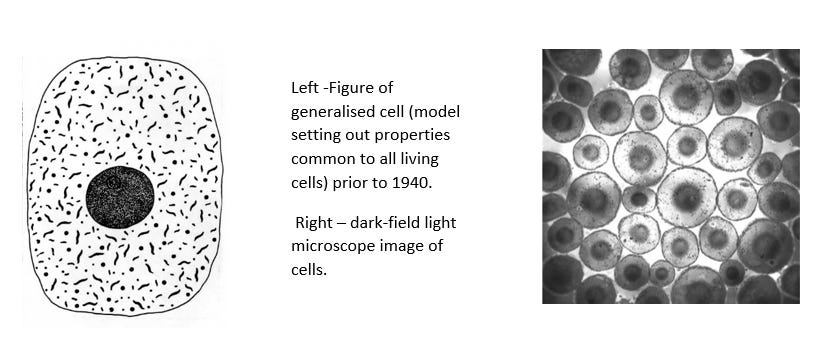
Following the introduction of the electron microscope, the following organelles were added to generalised cell and can be demonstrated to be artefacts (Source 5):
- The cell membrane, the nuclear membrane, and the mitochondria membrane now all appear as two lines with spaces in between them, this appearance is called the ‘unit membrane’. This is opposed to the single line accepted prior to 1940.
- An “endoplasmic reticulum” (ER) permeating the cytoplasm (gel like liquid inside cell membrane but external to nucleus) three dimensionally.
- “Ribosomes” line the endoplasmic reticulum.
- “Nuclear pores” appear in the nuclear membrane.
- “Lysosomes” a membrane bound cell structure found in the cytoplasm that contains digestive enzymes.
- “Cristae” were described in mitochondria.

The “Unit membrane”
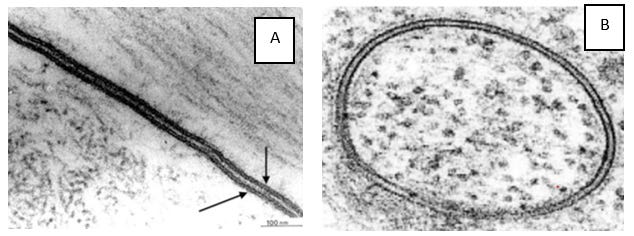
Reference: Living Cell pg 35 – 43 (Source 6)
The membranes of the cell, nucleus and mitochondria all appear on electron microscopy as two lines with a space in between, the latter sandwich been given the name “unit membrane”. See electron micrographs A and B above.
Hillman and Sartory point out several issues with the “unit membrane’s” deduced structure, below are the most obvious issues.
Geometric arguments
When you look at electron micrographs depicting membranes, the two outer lines of the sandwich always appear remarkably uniformly distant apart.
The problem here is that when you cut sections of concentric spheres (think layers of perfectly round onion), one expects to see that the apparent spacing of the layers will vary depending on how close the sections are cut to the center or the edge of the sphere (see figure 1).
Given that all membraned structures in a specimen will be randomly oriented, and that the microscopist can’t know in advance from which angle the microtome will cut, it follows that, even if the different layers had exactly the same thickness, they should appear in a variety of thicknesses in a series of sections. See figure 1.
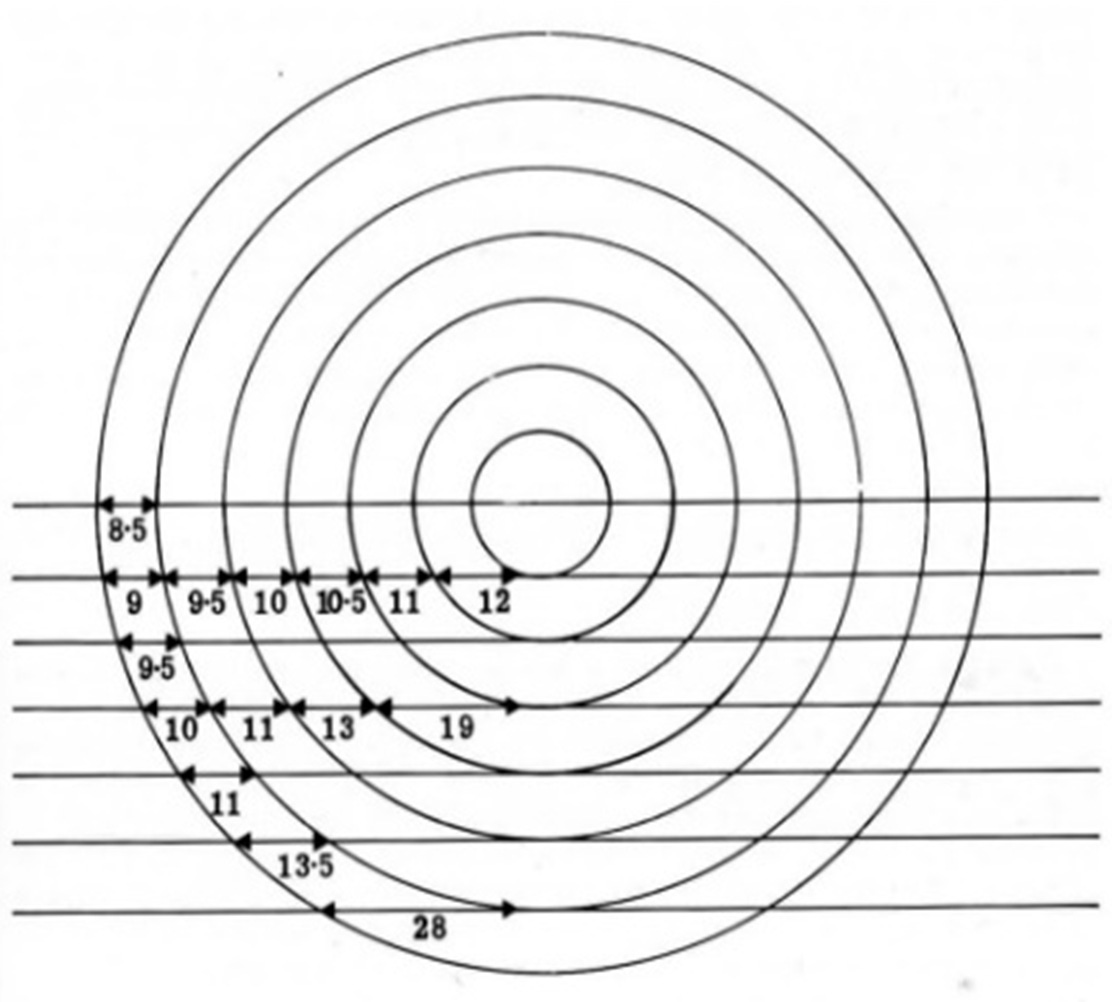
Figure 1.
In other words, in different sections taken of the same membrane, the thickness of the layers should vary and the amount of layers visible should also vary, in that there should be instances where only half of the layers are visible and others where the gap in the middle is not visible at all. See figure 2.
This is because how the layers of the membranes appear must necessarily depend upon where the section was made. The options being through the centre of the cell (three equal distant layers visible), through only a segment of the cell, close to the periphery of the cell, in between the two layers of a membrane (one layer and a portion of the middle gap visible) or entirely to the side missing the membrane (no visible gap).
The fact that on electron micrographs membrane layers are seen to consistently maintain their width no matter the orientation, means that all the structures visible in a particular section had to have been:
a) In a position that was perfectly central and symmetrical to each other at the time the section was made, and
b) Sliced exactly through the centre of their structures at the same time.
This is despite all visible structures varying greatly in size and position.
Such a series of interconnected circumstances is absurd and very unlikely. What is far more likely to be the case, is that the “unit” membrane is two dimensional and the double layer (suggesting three dimensions) appears either after the section is cut due to the preparation of the sample for viewing or during the electron bombardment itself.
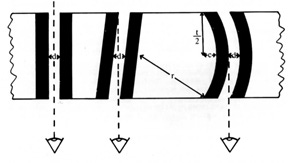
Figure 2:
A further problem with the appearance of “unit” membranes is the fact that the cell, nuclear and mitochondria membranes are all believed to be curved given that they encircle things. This means in any given single section we necessarily should be able to find instances where the space between the layers of a membrane is not visible due to the two outer layers appearing to overlap and cover the gap – as the double layers disappear around the corner, so to speak.
Instead, often we are presented with membrane layers that maintain their distance around the entire circumference.
Biological arguments and subcellular fractionation
When it comes to the biochemistry, it is generally believed that the “unit membrane” layers are two protein layers sandwiching a lipid (fatty) layer (Source 7).
At this point it is important to understand what subcellular fractionation is and how biochemists and molecular biologist use it to determine the location of various macromolecules (chemistry) within the living cell and examine the macrostructures (morphology) of the individual organelles.
Subcellular fraction is the process devised to separate cells into their different components (organelles) while leaving the organelles largely intact and preserving their individual functions.
It is important to note that it is believed that the procedures do not alter the properties of the organelles. That following their separation from the cell and each other, the organelles retain their properties to the same extent as if they were still in the intact animal.
The first step in subcellular fractionation requires the sample to be homogenised (ground, minced, chopped or pressured) within a non-biological buffer solution (a water based solution of reciprocating acid and basic substances).
The homogenisation ensures the cell membrane breaks and the cellular contents are released into the buffer solution. The buffer solution protects the organelles by maintaining the desired PH level of the solution and thereby inhibits enzyme activity (stops decay and chemical reactions from taking place). Refer to Source 8.
Sucrose is then added to the cell homogenate and then centrifuged. Centrifugation consists of putting samples in a centrifuge – a machine designed to spin liquid samples at high speeds – so that centrifugal force causes the larger or heavy particles sediment to form a pellet at the bottom of the centrifuge tube and the smaller or lighter particles to the top.
Sucrose in solution is used as a medium because its density is unaffected by centrifugation, so it does not interfere with the separation of organelles into sedimentary layers. A video of the centrifugation process can be reviewed by following – see source 9.
The speed, duration, and number of times a sample is centrifuged depends on which organelles are sought to be studied.
Generally, it is believed that the smaller organelles will require more rounds of centrifugation than larger organelles, and each round needs to be at higher speeds and for longer durations.
A low speed subjects the homogenate to a force 1000 times that of gravity for 10 minutes and a very high-speed subjects the homogenate to a force 150 000 times that of gravity for 3 hours (Source 10).

The sediments, known as fractions, which are pushed to the bottom after each round of centrifugation are then observed under electron microscope to determine their purity. If the fraction is determined not pure enough (there is more cellular debris and other organelles than the desired organelle), the fraction can be centrifuged again.
The macromolecules, such as proteins or nucleic acids, found in each fraction are then attributed to the organelles that should have separated into those densities. Note these macromolecules are not only attributed to the fractions but also to the organelles believed to be in the living cell.
Hillman and Sartory doubt that membranes are composed of lipids and proteins as they regard the evidence from the analysis of the chemistry of subcellular fractions to be too uncertain until the relevant controls have been done. Without controls being performed it is unclear what effect the processes of homogenisation, centrifugation and the necessary additives have on the sample and its biochemistry.
The entire procedure also rests upon the assumption that each organelle only contributes in one specific way to the biochemical processes of a cell (compartmentation). Refer to Source 11.
What is also noteworthy is that while mitochondria and nuclei are fairly easy to identify within fractions whereas membranes, ribosomes and the endoplasmic reticulum are not.
This is because ribosomes are without a clearly defined shape or form and the endoplasmic reticulum and membranes are said to be appear as circles or ovals in fractions. The circular particles seen in ER, ribosome and membrane fractions are known as “microsomes”. See electron micrograph C and Figure 3.

Figure 3: left diagram of ER and ribosomes under electron micrographs. Right diagram of ER and ribosomes after subcellular fractionation.
This is also important if one considers that the justification in subcellular fractionation for each manoeuvre and its duration, each additive and its concentration, is solely based on the apparent matching by electron microscope of structures in the fraction to those in the whole tissue.
A further reason why Hillman and Sartory doubt that membranes are composed of lipids and proteins is because during the fixation of tissues solvents (ethanol, acetone) are used which are known to dissolve lipids and as such it is difficult to believe that lipids are visible on an electron micrograph (Source 12).
Note the above issues concerning the appearance and biochemistry of the “unit membrane” are applicable to the endoplasmic reticulum discussed immediately below.
The “Endoplasmic reticulum” (ER)
The ER is regarded by many as one of the most important findings of electron microscopy (Source 13). Diagrams of it are found in all life science textbooks used by school children as well as by university students, postgraduates, and career research workers.
It is the largest organelle in the cell and is said to be a major site of protein synthesis, protein transport and protein folding (Source 14).
It is believed to be found in all plant and animal cells as a three-dimensional net or systems of flattened sacs or vesicles which permeate part or the whole volume of the cytoplasm. It also appears to have two layers with a space in between them as in the case of the “unit” membrane. See electron micrographs D and E below.

It is most often represented as being attached to the cell membrane and the nuclear membrane (Source 15). Sometimes the ER appears to be lined by “ribosomes” or granules, which are believed to be composed largely of ribonucleic acid (RNA) and also involved in the synthesis of proteins.
The ER is obviously believed to exist in living cells as they appear in electron micrographs, as implied in the view that it and the “ribosomes” which line it are involved in the synthesis of protein.
Not only has the existence of the ER been accepted universally among morphologists, but biochemists have produced subcellular fractions, which they identify as being from the ER (known as “microsomes” as explained above) and have studied its biochemistry extensively. Its function has thus been correlated with its structure, which is regarded as confirmatory evidence for its existence.
Hillman and Sartory’s biggest issue with the above explanation is that, in every electron micrograph provided as evidence of the ER in cells, the ER only ever appears in the plane of the picture in transverse section.
The problem with that is, if the ER were either a net or a series of flattened sacs, one would expect to see it in several orientations and not just one. A section would cut random portions of the ER if it were randomly orientated in the cell. Further, a series of sections should show a series of orientations.
In other words, if the ER is a net like thing made up of unit membranes why do we only ever see one orientation of it in cells? Where are the picture of all the different orientation that we would expect to see? It is quite impossible to conceive of a 3-dimensional object, which can always have the same appearance in two dimensions when sections are made of it in any orientation.
For example, why do we only see the ER on electron micrographs as shown in Figure 4a and 4b.

And never as shown in Figure 5a to 5f.

In response, electron microscopists have said that you do not see the other orientations of the ER because authors choose pictures in which you can see the ER clearly.
Hillman and Sartory say nonsense, not only had they themselves examined cells under the electron microscope but their investigations into these anomalies, lead them to examining electron micrographs of as many whole cells as they could find, thousands of unpublished and published electron micrographs of cells. Including ones in which the authors were drawing attention to structures other than the ER.
It is also noteworthy that – in complete contrast to the ER – mitochondria do appear in electron micrographs at every orientation from the cylinder to the circle, depending on the plane of the section. These can be seen in any randomly selected electron micrographs, and the image of these structures represents an excellent control model for observations on the ER.
See electron micrograph F below for example, the small dark circles and ovals of various sizes are the mitochondria in different orientations within the section. Of course, mitochondria can also be seen by a light microscope and their existence is not at issue.

A second reason why the ER appears to be an artefact is the fact that, in micrographs, the ER looks to be thicker than the cell and nuclear membranes, yet the latter two membranes can be detected with a light microscope. In addition, the current view that “ribosomes” line the ER in some cells would make the appearance of the ER thicker. Why is the ER not then detectable with the light microscope?
A further difficulty about the existence of the ER in the living cell concerns is its attachment to the cell and nucleus membrane. How can nuclear rotation occur if the ER is moored to both the cell and nucleus membrane, which is supposedly crossed with pores?
Nuclear rotation refers to the fact that under a light microscope you can observe random thickenings (bumps) along the nuclear membrane which rotate with the nucleus. This along with other intercellular movements has been observed in live cells since 1954 and is one of the criteria used by microscopists to tell whether the cells in the tissue sample are still alive.
The point here is, if the ER is attached to both membranes and one of them, the nucleus membrane, is constantly rotating a) how is the ER itself not becoming tangled, b) how are the mitochondria, granules and cell debris not getting caught up in the ER and c) how do particles introduced into the cytoplasm diffuse evenly and in all directions as they are seen to do under a light microscope?
If one were to argue that the ER is not attached to the rotating nucleus, it must then be admitted that the ER could not then act as the channel for exporting the proteins believed to be synthesised in the “ribosomes”.
So, what are electron microscopists seeing if not an ER? Hillman and Sartory have hypothesised that the ER is in fact merely the precipitation of the cytoplasm by fixatives.
Since living tissue is 60% water, dehydrating the tissue sample will cause significant shrinkage even if the water is subsequently replaced by alcohols, organic reagents and embedding media (epoxy resin or wax). The appearance of the ER is thus a result of the intense heat of the electron beam dehydrating and precipitating the metal-tissue combination. Whether this is correct or not, however, does not affect the conclusion that the ER does not exist in the living call.
“Ribosomes”
“Ribosomes” is the name given to granules seen on electron micrographs between the layers of the ER (see electron micrograph G), and to a particular subcellular fraction containing roughly 50% RNA (Source 16).

It is generally believed that RNA and its chemical function sits in the “ribosomes” in the living cell.
Hillman and Sartory argue that if the ER is an artefact due to deposit of the cytoplasm, it is very likely that “ribosomes” are also a deposit.
“Ribosomes” are not seen by light microscopy and Hillman and Sartory hold the view that, in addition to the issues they have with subcellular fractionation already discussed, there is insufficient evidence of subcellular localisation of biochemical activities. In other words, there is insufficient evidence demonstrating that the RNA detected after subcellular fractionation comes from the “ribosomes” in the living cell.
This is because all membranes are believed to be ‘rounded off’ into particles called “microsomes” and it is quite possible that “microsomes” in a particular fraction believed to be “ribosomes” on the ER could in actual fact be pieces of the cell membrane or any other membraned structure believed to be in the cell.
Further and given that it is believed that nucleic acid absorbs UV-light and thus can be used to detect nucleic acid, (Source 17) Hillman points out that if RNA were lining the ER, one should see this under ultra-violet microscopy as dark channels permeating the cytoplasm.
Put differently, where the ER and “ribosomes” are believed to be in the cell, that part should absorb much more UV-light and appear darker than its surrounds due to the high concentration of RNA. Hillman and Sartory could find no evidence that such experiments have ever been done.
“Nuclear pores”
Nuclei (the dark centre of cells) have been seen for 150 years (Source 18). Only recently, however, have “nuclear pores” been reported in nuclear membranes of many plant and animal cells by electron microscopy, though not by light microscopy.
A pore appears on transverse (crosswise) section of the nuclear membrane as a discontinuity in the two-line appearance of the membrane layers (see electron micrograph H). On a tangential, view as a circle or octagon (see electron micrograph I).

Initially, “nuclear pores” were thought to be breaks in the membrane but recently much more detail has been attributed to them.
Nowadays, they are said to be part of a “nuclear pore complex” and allow for the passage of proteins and RNA between the nucleus and cytoplasm, which is of particular importance to protein synthesis (Source 19 and Source 20).
As you can by now imagine, Hillman and Sartory have similar issues with this description as with the previous structures, namely the evidence presented appears to defy the rules of logic, geometry, and biology.
First, you only ever see “pores” as gaps in the “unit” membrane or as an array of circles or octagons, they are rarely, if ever, seen as any of the intermediary shapes which would ordinarily be expected. For example, the closer the “pores” get to the edge of the nucleus they should necessarily appear more squished or slit like, rather than maintain their appearance right up to the edge (See figure 6).

Figure 6: Nucleus pores.
Second, it must be realised that you will only be able to see gaps in the “unit membrane” in instances where the “pore’s” diameter is thicker than the section thickness and the section cuts the “pore” on both sides (front and back faces). See figure 7 (b). Note it follows that the pore will necessarily appear to have the diameter of the hole of the smallest face cut by the section and will not always appear to be of the same diameter in every section.
Third, similar circles or octagons with similar dimension appear in the cytoplasm but these are never deemed to be pores. Indeed, the criterion of a circle of relatively uniform in diameter on the nucleus appears to be the only way of identifying “nuclear pores”.

Fourth, even if “nuclear pores” are said to only perforate 3 percent of the nucleus membrane it is difficult to understand how nuclei apparently maintain their approximate spherical shape after homogenisation or centrifugation.
Maintaining a spherical shape would require either a consistent pressure difference between the inside and outside of the nucleus – which would not be possible if the membrane were punctured with pores – or an internal structure maintaining its spherical shape.
Fifth, if “nuclear pores” allow RNA to pass through, how do they prevent smaller molecules and ions from also passing through, and then how can it be said that there is a potential difference (a difference in PH) across the nuclear membrane? (Source 21).
In other words, how is the difference in PH (difference in hydrogen ions) between the nucleus and cytoplasm maintained if the pores allow elephants (mRNA molecules) to pass through but not mosquitoes (hydrogen ions)?
Hillman and Sartory argue that it is far more likely that “nuclear pores” are cracks in the nucleus membrane occurring during the fixation, dehydration, freezing, or subjection to the electron microbeam.
An accurate model of the living cell
Note that “lysosomes”, “Golgi apparatus” both said to be found in the cytoplasm and “cristae” said to be located in mitochondria are also organelles discovered by electron microscopy and said to exist in the living cell. They are not, however, discussed in this paper as they are not directly relevant. Nevertheless, the reader should know that these organelles have also been shown by Hillman and Sartory to be artefacts, for similar reasons as set out above, namely they appear to defy the rules of logic, geometry, and biology (Source 22).
Ultimately, Hillman and Sartory regard the evidence for the cell membrane, the nucleus, the mitochondria, cytoplasm and nucleus existing in the living cell as beyond reasonable doubt. The rest of the currently accepted organelles of the living cell seem to be far from certain.
One more time for those of you who are not concentrating, save for the nucleus and mitochondria, all other organelles said to be found in plant and animals cells are artefacts produced by the electron microscope and do not exist outside electron micrographs. The pre-1940 generalised cell model is the only accurate model of a living cell.
Protein synthesis, the central dogma of genetics and DNA
As alluded to in the above discussion of Hillman’s work, the main function attributed to the organelles of the cell “discovered” by electron microscope relates to the manufacturing of proteins. How protein synthesis became synonymous with gene expression and the central dogma of genetics will be explained in further detail shortly, it is first necessary, however, to briefly look at how molecular biologist determined that protein synthesis takes place in cells. It is also noteworthy, that it is believed that the living cell is primarily comprised of water, proteins and potassium which floats in an environment of water and sodium.
Cell free protein synthesis (CFPS)
The mechanisms said to be responsible for protein synthesis were discovered by biochemists and molecular biologist in the 1950s and 1960s, two of the most crucial aspects being the discovery of RNA (tRNA and mRNA) and the realisation that RNA was located in the ER and “ribosomes” of all cells.
It was these crucial discoveries that lead biochemists and molecular biologists to being able to synthesise proteins in test tubes by incubating “ribosomal” fractions (RNA), also known as “microsomes”, with certain substrate mixtures – amino acids (building blocks of proteins), salts (sodium and potassium to replicate environment of living cells), adenosine triphosphate (ATP’s).
ATP is macromolecule which is said to be created by the mitochondria in cells out of the glucose derived from the food we consume (process is known as glycolysis). It is said to function as a high-energy storage molecule used in the cell. When the bonds of the ATP molecule are broken via acid hydrolysis in the “lysosomes”, energy is released which is harnessed by the “ribosomes” to synthesis proteins. It thus believed to be the energy source powering protein synthesis (Source 23).
Basically, the mechanisms of protein synthesis were deciphered in experiments which began with a cell homogenate that could synthesis proteins. Subcellular fractionation of the cell homogenate allowed individual fractions to be added or withheld separately from the incubated substrate mixture to determine its exact role in the process and which combinations lead to the amino acids in the tube being synthesised into proteins.
This procedure is known as ‘cell free protein syntheses’ (CFPS) and forms the basis for synthetic biology. The subcellular fraction necessary for protein synthesis to take place in the test tubes were then later identified by means of electron microscopy to be derived from the ER and “ribosomes”.
Whether CFPS demonstrates beyond doubt that cells synthesise proteins, is a topic for another paper. But given that that the underpinning experiments were only possible using subcellular fractionation, an electron microscope, and chemical treatments none of which have been demonstrated through control experiments not to interfere with the experiments themselves (as discussed in this paper already), it is fair to say that these discoveries rest on rocky foundations.
For now, though, we will give them the benefit of the doubt, and accept for purposes of this paper, that cells do synthesise proteins. What is not accepted, however, and as evidenced by Hillman’s work, is that proteins are manufactured by the ER and the “ribosomes” in cells.
For those wondering, proteins are said to perform a wide range of crucial functions in living organisms. They act as enzymes, hormones, antibodies, structural components, and transport molecules. Protein synthesis allows cells to produce these necessary components in response to changing needs, such as growth, repair, and regulating physiological processes. Basically, proteins determine how your body is structured and how it functions.
Gene expression and the central dogma
Gene expression forms the central dogma of molecular biology and genetics and, since Francis Crick first proposed it over fifty years ago, the central dogma model has come to dominate research in these fields. Simply put, the central dogma is the origin story for RNA, without which (cell free) protein synthesis could not take place.

The central dogma states that DNA is static, it does not change of its own accord from the moment it comes into existence; from DNA comes RNA (transcription); RNA is translated into proteins by the protein manufacturing machinery in cells (translation). This process can only flow one way and proteins cannot be made to transform back into genetic material. Lastly, because DNA is made of the same four nucleotides in every living thing, the genetic code is universal, which means that a gene should be capable of producing its particular protein wherever it happens to find itself, even in a different species.
Protein synthesis, carried out by the ER and “ribosomes”, is thus believed to be under the exclusive control of our DNA and the genes from which it is comprised. This is because it is believed that the sequence of the nucleotides in a gene “is a simple code (recipe) for the amino acid sequence of a particular protein (Source 24).
To briefly explain the gene expression process: An enzyme (a type of protein), known as RNA polymerase, is responsible for the transcription of a gene (a segment of DNA) into what is called messenger RNA (mRNA).
Following its creation, mRNA leaves the nucleus for the cytoplasm via the “nuclear pores” and floats its way to the protein making machinery of the cell. As you by now know, the “endoplasmic reticulum” and particularly the “ribosomes” are the organelle said to be responsible for protein synthesis and thus for translating the mRNA and assembling amino acids into protein chains (peptides) in accordance with the recipe imparted by the mRNA molecule.
A molecule known as transfer RNA (tRNA) is thought to be attached to the amino acids and functions as the “ribosomes” translator or adapter. This is because the “ribosomes” are said to read the mRNA recipe three nucleotide base pair (codons) at a time and then match them with the complementary anticodons on the tRNA molecule (Source 25).

It is important to note that no person has ever observed the central dogma in action in any living cell. All the structures and molecules claimed to be involved cannot be seen under a light microscope in unfixed tissue (live tissue). The structures are observed only under electron microscope and the molecules found only after subcellular fractionation.
The central dogma is therefore a theoretical model proposed after observations were made of a) tissues that had undergone various procedures and chemical treatments and b) static pictures obtained from electron microscopes. Moreover, it is very hard to believe this process is supposed to transpire in exactly the same way in a test tube following subcellular fractionation.
DNA
DNA, where RNA is supposedly derived, is said to be a very long, linear molecule shaped in a double-helix and tightly coiled within each cell’s nucleus.
DNA is believed to be made up of four different kinds of nucleotides, strung together in each gene in a particular linear order or sequence. The four kinds of nucleotides can be arranged in numerous possible sequences, and the choice of any one of them in the makeup of a particular gene represents its “genetic information” (Source 24).
Each human cell is believed to contain roughly 2 metres of DNA and is supposed to be packed into a nucleus of 6 nm. This is akin to packing 40 km of extremely fine thread into a tennis ball.
Apparently, this is possible because four double-stranded DNA strings are wrapped and tightly folded around a histone molecule (type of protein), into packages known as a nucleosome. Nucleosomes are believed to be condensed further into a spiral structure known as chromatins, which are finally condensed further into chromosomes.

It is claimed that humans have 46 chromosomes which exist in pairs, one form our father and one from our mother, and that within our nuclei there are 23 paired chromosomes in a roughly linear rod-like structures. The total sequence of these strings of DNA within these chromosomes are then known as the human genome (blueprint). See Source 24.
Unbelievably, despite being very tightly folded and wrapped up and condensed into complex structures there are apparently no issues when it is necessary for the strands to be copied and transcribed in mRNA (Source 26).
It is essential to note, that molecular biologists feel qualified to make these assertions about DNA’s composition and structure because they claim to have isolated the DNA found in cells. In many respects these claims are similar to those made about the inner structure of cells in that they fail to take into account the impact the experiments, which led to the discovery, had on the actual things said to be discovered.
I cannot critically review these claims any better than has already been done by Tam in her paper on the subject, (Source 27) I would suggest that Tam’s paper is mandatory reading for anyone wanting to know about DNA.
The bottom line is what molecular biologists have isolated cannot honestly be said to be anything more than the by-products that remain after a biological sample has been mixed with chemicals and then heated. This is especially the case since no control experiments have been performed. If the isolation of DNA is dubious it follows that the structure of the molecule is equally suspect.
Central dogma and heredity
It’s crucial to realise that the central dogma reduces the creation of life and inheritance to chemical reactions in molecular dimensions, this is because DNA has been pointed to as the molecular agent of inheritance.
What lead to this conclusion is the fact that when observing cells dividing (mitosis) under a light microscope, it is possible to see stringy bits in the nucleus of a cell divide and then separate with one half remaining in the original cell and the other moving into the nucleus of the newly formed daughter cell. It was then proposed that these stringy bits were chromosomes and the reason all future cells produced identical proteins.
Note, Hillman pointed out that this is only observable in fetal cells and chromosomes cannot be seen in all living cells (Source 28). Fetal cells are only found in embryos or unborn babies.
DNA (packaged into chromosomes) is therefore believed to be the recipe book of how to assemble a particular human. The recipes (our genes) in the book are a combination of recipes believed to derive from the recipe books of either our farther or mother. In other words, our cells can only produce proteins in accordance with recipes received from either of our parents.
This means if our parents, or anyone else in our lineage, has faulty or unique recipes there is a chance that these recipes will be passed onto us and our cells will manufacture defective or unique proteins, which in turn will have a particular impact on the structure or function of our body.
Concluding remarks
When one overlays Hillman’s work concerning the living cell onto the theory of the central dogma and protein synthesis as currently accepted it becomes obvious that these models cannot work as believed. The structures said to be involved in the process and which give credibility to the theory, simply do not exist.
When Hillman tried to point this out to the scientific community, he received mostly hostility and all the “reputable” and well-known scientific journals refused to publish his papers on the topic. As a result, Hillman and his work has been completely ignored and buried under mountains of inexact “science”.
Further issues with protein synthesis and the central dogma
In addition to the fact that the “endoplasmic reticulum” and “ribosomes” do not exist in the living cell, it is important to be aware of a few further crucial issues Hillman pointed out with the current accepted model of protein synthesis.
First, where does protein synthesis take place in cells which have no “ribosomes”? See Source 29.
Two, what is the evidence for the claim that each cell of a particular plant or animal contains the exact same quantity of DNA, especially since there is a huge variation in the size of nuclei of cells? See Source 30.
If it can be shown that cells from different parts of our body have different amounts of DNA – as shown in these articles discussing recent studies to this effect (Source 31) – the central dogma (one gene equals one protein) and its accomplice, surely protein synthesis via “ribosomes”, can no longer be held up as irrefutable fact?
Third, until such time as the proper controls have been carried out for subcellular fractionation it cannot be said irrefutably that DNA is located in the nuclei of cells (Source 32). Note further that DNA and RNA is only ever extracted from dead tissues.
Hillman did not make the next few points, but they are nevertheless relevant to this discussion.
Fourth, if the central dogma is correct, it follows that in each living thing there should be a one-to-one correspondence between the total number of genes and the total number of proteins. The entire array of human genes, that is the genome, must therefore represent the whole of a person’s inheritance or blueprint, which distinguishes a person from a fly.
In the 1990s the Human Genome Project set out to identify and enumerate the entire human genome by working out the sequence of the 3 billion nucleotides in human DNA. However, to their surprise their findings contradicted the central hypothesis that one gene equals one protein. Instead of finding the 100 000 or more genes predicated by the estimated number of human proteins, they only managed to find about 30 000 genes.
To put this in perspective, this would make humans as genetically rich as a mustard weed plant which has 26 000 genes and twice as genetically endowed as a fruit fly (Source 24). As such our “genes” cannot explain the vast inherited difference between a weed and a person. This is also apparent if one considers the fact that humans, who are considered to be the most complex of Earth’s creatures, have less genes than animals and plants (Source 33).
Fifth, it appears that the claim that DNA is static was disproven decades ago by Barbara McClintock, when she described how “extragenic” components, known as transposons, could alter the function of nearby genes (Source 34). Meaning that the mechanism controlling genes could be independent of the genes themselves. Ironically, McClintock’s conclusions about DNA instability were published the same year as reports describing the double-helix structure of DNA by Francis Crick and James Watson were published.
Sixth, it appears from recent studies that biochemists are still finding new nucleotide base pairs, indeed it is claimed that in fact there may be seven or eight base pairs (Source 35 and Source 36). This completely defies our understanding of the structure of DNA and its role in the central dogma of genetics.
Where does this leave us?
Hopefully from the discussion of the accepted science above, it is clear that:
- Living cells do not appear in nature as they do in electron micrographs.
- Save for mitochondria and the nucleus, there are no organelles within the living cell.
- The ER and “ribosomes” do not manufacture proteins in accordance with mRNA molecules fashioned in accordance with our genes and DNA.
- Our genes are not recipes for a single protein.
- The central dogma of genetics needs to be scrapped.
- The entire concept of DNA needs to be reappraised.
What are some of the everyday implications of these revelations?
First, it should be abundantly clear by now that genes cannot account for our inherited traits. It has not been shown that DNA is ultimately in control of protein synthesis, nor has it been demonstrated that DNA is static or consistent in every cell.
This does not only mean that genes do not determine our hair and eye colour etc but it also means they cannot be responsible for our susceptibility to certain “diseases” – for example autoimmune diseases and psychiatric illness.
It also means that cloning is utter nonsense because there is no human genome to be replicated and assembled and even if there were we have no clue as to how proteins are created. Please note I am note denying that inheritance of traits happens, rather I am simply stating we have no idea as to how it happens.
Seeing as viruses are all the rage lately let us do those next. Viruses are said to be pieces of genetic material wrapped in a protein coat which act as intracellular parasites that hijack the cell’s protein machinery and pathways (ribosomes and endoplasmic reticulum) in order to replicate.
Basically, whether they are said to be RNA or DNA viruses the process is the same. The virus makes its way into the cell and inserts its genetic material into a cell’s protein manufacturing machinery so that the cell produces the viral proteins necessary for the virus to replicate, instead of producing the proteins required by the living organism. Well, if the protein machinery does not exist exactly how does replication, then take place?
If you combine this with the lack of isolation of a virus (Source 37 and Source 38) and the numerous failed transmission studies that have taken place over the last 150 years (Source 39), viruses seem to be nothing but the fairy tale of virologists and all vaccination a sick joke.
Another causality of this pseudoscience is the theoretical basis of the multibillion-dollar industry of genetically modified food crops. When genetically engineering plants, biologist claim that they insert beneficial genes into the DNA of a plant thereby providing that plant with the recipe or code necessary to make proteins which will allow them to be immune to the effects of certain insecticides and herbicides.
However, it is clear from the above discussion that:
a) Cells do not have protein factories.
b) Genes have not been demonstrated to be in control of protein synthesis.
c) Even if genes as described do exist, they are more likely to be responsible for the creation of many different types of proteins and not just one.
Gene-therapy is pretty much just an attempt to genetically modify humans instead of food crops and as such runs into the same three issues set out above with GMO. For example, what about the vaccines that are said contain mRNA and could result in gene- therapy?
It is claimed that the purpose of mRNA in certain vaccines is to instruct our cells to make a specific protein, a viral spike protein, which will cause our bodies in turn to then make the appropriate antibodies which will defend the body against a specific virus. Gene-therapy on the other hand involves making deliberate changes to a person’s DNA by introducing a gene which allows the cells to make a specific protein that cures or alleviates a genetic condition.
Gene-therapy and mRNA vaccines converge because it is feared that if the mRNA from the vaccines is not converted into the spike proteins it may then wonder into the nucleus via the “nuclear pores” and join with our very tightly wrapped DNA, thereby permanently altering our DNA and all its daughter cells from there onwards. Again, if cells do not have protein factories and “nuclear pores” do not exist how is this all supposed take place?
Finally, it must be said that all scientific research which takes for granted the structure of cells, DNA and the process of protein synthesis, like virology and genetics, is a gigantic waste of time. This includes all the time spent studying these subjects in university. These two branches of medical science are nothing more than ‘brain drains’ in that they literally drain people away from focusing on subjects of substance.
Conclusion
If you finish this paper only having learned one thing, let it be the fact that the vast majority of the work and research of biochemists, molecular biologists and electron microscopists (i.e. those people who talk about DNA, RNA, genes and molecules) carried out over the last 100 years is not infallible science and is not trustworthy. Whether they have fooled themselves or simply do not practice intellectual honesty, it is not knowledge that our health and every day lives should be based on.
The work of these scientist has done nothing but incite fear and convey intellectual authority to people who are not worthy, and it needs to stop. Humanity is becoming more and more enslaved by its own stupidity and materialism.
The only way out is to take back knowledge from the hands of a few specialists and return it into the hands of the majority. The way to do this is to make things understandable for the majority and not over complicated and incomprehensible. It is my hope that this paper is a step in that direction
Subscribe to dpl Newsletter
Reference List:
Source 1 – Harold Hillman Obituary
Source 2 – Harold Hillman Biology
Source 3 – Hillman, H. (1972). Certainty and Uncertainty in Biochemical Techniques. Surrey University Press, Henley on Thames.
Source 4 – Tom Cowan podcast on Harold Hillman’s 47 Questions (start at 03:26)
Source 5 – Hillman, H. and Sartory, P. (1980). The Living Cell. Packard Publishing Company, Chichester (“Living Cell”) – Chapter 1
Source 6 – Living Cell pg 35 to 43
Source 7 – The Cell: A Molecular Approach. 2nd edition.
Source 8 – Hillman, H. (2008). Evidence-based Cell Biology, with some Implications for Clinical Research.
Source 9 – Centrifugation.
Source 10 – Fractionation of cells.
Source 11 – Certainty and Uncertainty in Biochemical Techniques, Harold Hillman Surrey University Press, 1972. See full list of 23 assumptions underlying subcellular fractionation.
Source 12 – Harold Hillman 47 unanswered questions in biology – See question 12.
Source 13 – Living Cell pgs 43 – 60
Source 14 – Molecular Biology of the Cell. 4th edition.
Source 15 – Amber R. English, 2013. Endoplasmic Reticulum Structure and Interconnections with Other Organelles.
Source 16 – Living Cell pgs 60 -61
Source 17 – Benjamin J Zeskind, 2007. Nucleic acid and protein mass mapping by live-cell deep-ultraviolet microscopy
Source 18 – Living Cell pgs 67 -75
Source 19 – Molecular Biology of the Cell. 4th edition.
Source 20 – The Cell: A Molecular Approach. 2nd edition.
Source 21 – Harold Hillman 47 unanswered questions in biology emails see question 32
Source 22 – Living cell – lysosome pg 66-67; Golgi app pg 61 – 66, Cristae pg 61
Source 23 – How cells obtain energy.
Source 24 – Unraveling The DNA Myth: The Spurious Foundation of Genetic Engineering.
Source 25 – From RNA to protein.
Source 26 – Chromosomal DNA.
Source 27 – Critical Check DNA Discovery, Extraction and Structure. A Critical Review.
Source 28 – The Amino Age and The New abNormal Doctors.
Source 29 – Harold Hillman 47 unanswered questions in biology emails see question 7.
Source 30 – Harold Hillman 47 unanswered questions in biology emails see question 33
Source 31 – Surprising science: Not all our cells have the same DNA.
Source 32 – H Hillman, 2008. Evidenced Based Cell Biology, with some implications for clinical Research. pg 29
Source 33 – List of organisms by chromosome count.
Source 34 – Halpern ME. Barbara McClintock on defining the unstable genome. Genetics. 2016; 204:3-4.
Source 35 – Genetics: Biosynthesis pathway of a new DNA nucleobase elucidated.
Source 36 – Scientists identify seventh and eighth bases of DNA.
Source 37 – Tom Cowan Breaking the Spell
Source 38 – What Does—and Doesn’t— Make Us Sick.
Source 39 – Virology the damning evidence by dpl
Some additional sources that I read/watched in preparation for writing this article, and which are related:
- Hillman [1, 2, 3]
- Jamie on DNA and genetics
- Omar Jordan on DNA Fingerprinting
- Horius on DNA
- Dawn Lester on genetics




Thanks for sharing the article! ??